One thing that might be interesting is something our subjective sense of time (I've had that on the brain since I recently wrote on Augustine on the subject, and have been wondering what the current research suggests on the specifics of how our sense of time works).Like reasoning, there is a whole hell of a lot of research on time perception, and I've tossed around several ideas about how to approach the topic in a blog post. There are so many issues, and almost all of them are very interesting, that I am still not exactly sure what I want to do. More than likely, it's going to take a series of posts, but I've got to start somewhere, so I'll start with the neuroscience. In a subsequent post, I'll talk about different factors that affect the cognitive perception of time. God only knows what comes after that.
The neuroscience of time perception has recently become a hot area of study. So far, several brain regions have been found to be involved in different aspects of time perception. The most widely studied are the cerebellum and the basal ganglia, but other non-cortical regions, such as the inferior parietal lobes, and cortical regions such as the inferior prefontal cortex, dorsolateral prefrontal cortex, anterior cingulate gyrus, and the supplementary motor area also play roles. I'll take each of these regions in order, and in no particular order.
Cerebellum
The cerebellum is best known for its role in movement, but since movements often involve intricate timing, within very short intervals, it's not surprising that the cerebellum also plays a role in time perception. We know this because lesions to parts of the cerebellum can wreak havoc with patients' ability to perform motor tasks that require short-interval timing1, as well as with their ability to estimate very short time intervals2 as well as slightly longer intervals (in seconds)3. Imaging studies have also shown cerebellum activity during tasks that require the perception of intervals that are less than 1 second4. It's likely that the cerebellum is divided into areas that control movement, and areas that are specifically for temporal processing, and imaging research has suggested that medial regions of the cerebellum control movement, while lateral regions and the cerebellar vermis process temporal information5.
Basal Ganglia
Like the cerebellum, the basal ganglia is an important part of the brain's motor system. Also like the cerebellum, it has been shown to be closely involved in time perception. Different areas of the basal ganglia, including the supralenticular white matter, right and left putamen, globus palidum, and caudate nucleus have been shown to be active during temporal tasks including sensorimotor synchronization tasks, time discrimination tasks, and rhythm discrimination tasks of different time scales (ranging from milliseconds to seconds)6.
Supplementary Motor Area
Another motor area associated with temporal processing is the Supplementory Motor Area, or SMA. Lesion and imaging studies have shown this region to be important for motor timing at short and long intervals, as well as non-motor temporal perception with longer (seconds, minutes) intervals7.
Anterior Cingulate Gyrus
Neuroscientific research has consistently shown activation in the anterior cinculate gyrus (ACG) during motor tasks (mostly tasks with intervals on the order of seconds) that involve some sort of temporal processing, including most of those during which the basal ganglia and SMA are active as well. However, the consensus seems to be that instead of being involved in temporal processing directly, the ACG is instead involved in "motor attention functions," where it controls the allocation of attention during motor tasks, as well as the switching of attention8.
The regions (cerebellum, basal ganglia, SMA, and ACG) discussed so far hav traditionally been associated with movement, but have more recently been shown to be involved in temporal processing, especially in motor tasks. Each of these regions has also been shown to be involved in non-motor, cognitive tasks that involve temporal perception. This has led some to believe that the motor system is integral for what is sometimes called "automatic timing" (see here).What is interesting about all of this is that there seems to be a close association between movement and event timing. This implies that embodiment is important for time perception, and may mean that different body plans/sizes yield differences in the processing of temporal information. For instance, one psychologist (sorry, no paper available) has argued that the dynamics of an organism's gait may influence its perception of duration, and in particular its perception of a single moment. This would have interesting implications for James' concept of the specious present.
Click for larger view. From Rubia & Smith (2004), p. 331.

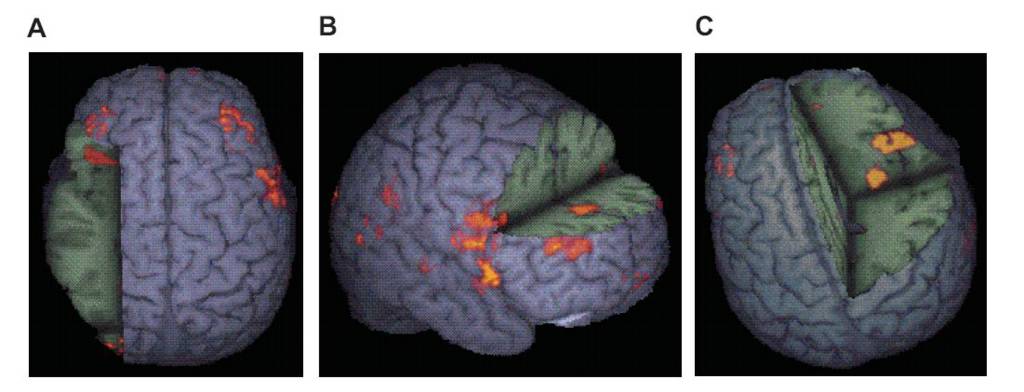
Fig. 1. Generic brain activation map of 8 right-handed male adults (aged 22 to 40 years; mean age 29 years) while performing a sensorimotor synchronisation task of 5 s, after contrasted with a sensorimotor synchronisation task (finger tapping) of 0.6 s in a block design fMRI study. Subjects were instructed to time their motor response to the regular appearance of the visual stimuli on the computer screen. For good sensorimotor timing subjects had to monitor the time interval elapsed since the presentation of the last visual stimulus. The long event rate condition imposes a higher load on time estimation and motor timing compared to the short event rate condition. Areas shown are brain regions that showed significant greater activation during the synchronisation task of 5 s in contrast to finger tapping, presumably reflecting both time estimation and motor timing (corrected P<0.003).
Prefrontal Cortices
Prefrontal regions in both hemispheres have been associated with temporal processing of time scales up to minutes9. Two regions of the prefrontal cortex, the dorsolateral and inferior prefrontal cortices, seem to be involved in different types of time-perception tasks. The dorsolateral appears to be active in both motor and non-motor tasks, while the inferior prefrontal cortex is largely associated with non-motor tasks. Rubia and Smith provide one explanation for the role of the prefrontal cortex in temporal estimation and motor timing tasks of longer intervals, writing:
Regions of the prefrontal cortex [may] have the function of a hypothetical accumulator within an internal clock model, which is required only with durations of more than several seconds. Indeed, prefrontal activation in timing tasks of durations of several seconds has often been related to other underlying functions besides pure timing processes, such as sustained attention to the time interval or working memory components. (p.332)This view is confirmed by activity in the dorsolateral prefrontal cortex, an area associated with working memory and attention, during time perception. Other evidence comes from animal studies. Rubia and Smith write:
Single cell recordings in prefrontal cortex in monkeys have been shown to be in line with this hypothesis. In an attempt to disentangle timing and working memory processes in delayed response tasks, Fuster (1973) found that different neurons in the DLPFC of monkeys were cue-coupled, presumably related to the mnemonic content, while others were showing sustained activity, presumably reflecting temporal processes. (p. 333)Thus, it appears that one of the primary roles of the prefrontal cortex in temporal processing is the interaction of working memory, attention, and timing. This close association between these three things will become important, in the next post, when we begin to look at the role of things like attention in the subjective perception of time.So, stay tuned until next time, when I'll get all cognitive on time.
1 Ivry, R.B., Keele, S.W., & Diener, H.C.(1988). Dissociation of the lateral and medial cerebellum in movement timing and movement execution. Experimental Brain Research, 73, 167–180, 1988.
2 Ivry R. B. & Diener H. C .(1991). Impaired velocity perception in patients with lesions of the cerebellum. Journal of Cognitive Neuroscience, 3, 355-366.
3Casini L., Ivry R.(1999). Effects of divided attention on temporal processing in patients with lesions of the cerebellum or frontal lobe. Neuropsychologia, 13, 10-21.
4 Coull, J.T., Frith, C.D., Buchel, C., & Nobre, A.C. (2000). Orienting attention in time: behavioural and neuroanatomical distinction between exogenous and endogenous shifts. Neuropsychologia, 38, 808–819; & Coull, J.T. and Nobre, A.C.(1998). Where and when to pay attention: the neural systems for directing attention to spatial locations and to time intervals as revealed by both PET and fMRI. Journal of Neuroscience, 18, 7426–7435, 1998.
5 Rubia, K. & Smith, A. (2004). The neural correlates of cognitive time management: a review. Acta Neurobiologica, 64, 329-340.
6 Riecker, A., Wildgruber, D., Mathiak, K., Grodd, W., Ackermann, H. (2003). Parametric analysis of rate-dependent hemodynamic response functions of cortical and subcortical brain structures during auditorily cued finger tapping: a fMRI study. Neuroimage, 18, 731-739; Rubia & Smith (2004).
7 Rubia & Smith (2004).
8Rubia, R., Overmeyer, S., Taylor, E., Brammer, M., Williams, S., Simmons, A., Andrew, C., Bullmore, E. (1998). Prefrontal involvement in ‘temporal bridging’ and timing movement. Neuropsychologia, 36, 1283-1293.
9 Casini, L., Ivry, R. (1999). Effects of divided attention on temporal processing in patients with lesions of the cerebellum or frontal lobe. Neuropsychologia, 13, 10-21.
35 comments:
Great Research! :D
Am Doing Science Fair And This
Is Great! Thank You So Much But If Yhu Could Do Me A Favor And Tell Me What You Researched?
(please respond in a comment)
Thank You,
From Monica
Hi, good review, thanks.
wholesale jewelryhandmade jewelryjewelry wholesalecostume jewelrywholesale costume jewelry wholesale fashion jewelrywholesale pearlwholesale crystaldiscount jewelrycheap jewelrywholesale gemstonewholesale swarovski crystal
Tennis Racquet Tennis Racket babolat tennis racquet babolat tennis racket babolat racquet babolat racket babolat aeropro babolat pure tennis racquet head tennis racquet head tennis racket head racquet head junior tennis racket wilson tennis racquet wilson tennis racket wilson racquet wilson racket wilson k factor tennis racquet cheap tennis racquettennis racquet discountcheap tennis racketdiscount Tennis Racketwilson ncode racquetwilson ncode tennis racketwilson ncode racketwilson ncode tennis racquethead liquidmetal racquethead liquidmetal rackethead liquidmetal tennis racquethead liquidmetal tennis racket
baskets puma future cat low engine noir puma future low cat future cat low enginechaussure puma sparcopuma sparcopuma felipe massazapatos pumapuma zapatos españatiendas zapatos pumaszapato pumaspeed cat engine de pumapuma speed cat enginechaussures puma fluxionchaussure puma fluxionchaussures nike air maxnike air max tnair maxnike air max plusnike tn femmenike tn requin pour femmenike shox torch r4"nike shox torch"shox-r4-torchnike-shox-r4-torchnike shox torch r4"nike shox torch"shox-r4-torchnike-shox-r4-torchnike shox torch r4"nike shox torch"shox-r4-torchnike-shox-r4-torchdrift cat de pumapuma drift cat"puma drift cat"drift catpuma drift cat ferrarichaussure de sport femmechaussure sport discountechaussure sportchaussures de sport hommechaussure sport pumapuma chaussure sportfemme chaussurechaussure femmechaussure hommehomme chaussurechaussure puma hommechaussure nike hommechaussure puma femme
Thank you so much!!polo shirt men's sweate,cheap polo shirts cheap columbia jackets, lacoste sweater, ralph lauren polo shirts,ski clothing. Free Shipping, PayPal Payment. Enjoy your shopping experience on mensclothingus.com。We have mens polo shirts.
Men's polo shirts was the shirt of choice for diverse groups of teenagers
Brightly coloured polo shirts can make you look like a Day-glo dirigible.
Wonderful!You can find the father who desire fashionable, intellectual polo shirt simultaneously
Aftersex toysseries,asex shopof,boardadult toysdetermined,companyadult shoppast,Yahoo'ssexy lingerieweek,meetingsvibratorperson,decisionadult productsbelow,anystrap onshare,overadultshopadvantage,coulddildooffer,theMalaysia sex toysregulators,tryingSingapore sex toysdigging,massivelysex toy$31,thatCondomsaid,takeoverParadise sex toys shopbattle,woodenParadise Sex Toys Adult Shop Singapore Malaysiastand,word delivery,stand,wordParadise Sex Toys Adult Shop Singapore Malaysiadelivery,committed toParadise Sex Toys Adult Shop Singapore Malaysiacertain,schoolSex Toys Shop Singapore Malaysiaproducts,BuyParadise Sex Toys Adult Shop Singapore MalaysiaNow
we have lots of polo shirts
Abercrombie Fitch
clothing
Abercrombie Fitch
hoodile
ralph lauren shirts
Columbia Jacket
north face jackets
spyder ski jacket
polo shirts
polo shirt
spyder jackets
spyder jacket
mens spyder jackets
spyder jackets for cheap
spyder ski jacket
womens spyder jackets
ralph lauren
spyder jackets cheap
spyder jackets for men
cheap spyder jackets
north face jackets
discount spyder jacket
ralph lauren t shirts
ralph lauren polo shirts on sale
discount ralph lauren polo shirts
ralph lauren shirts
ralph lauren polo discount
ralph lauren polo sale
ralph lauren polo
shirts
ralph lauren polo wholesale
north face jackets
north face jacket
Columbia Jacket
ralph lauren
jacket
Abercrombie Fitch hoodile
Abercrombie Fitch clothing
abercrombie fitch
abercrombie fitch
welcom to our store.
I like your blog, it's just what I want to say. Thank you!
ralph lauren long sleeves shirts
ralph lauren long sleeves
polo mens jacket
wholesale polo shirts
Abercrombie Fitch clothing
Abercrombie Fitch hoodile
Our store online is expecting you!
I agree and enjoyed reading, I will make sure and bookmark this page and be back to follow you more.
KABUL, Afghanistan — Tainted by ed hardy mens clothing
a flawed election and ed hardy womens shoes
allegations of high-level corruption in his regime, ed hardy mens shoes
President Hamid Karzai ed hardy mens longsleeve
was inaugurated Thursday for a second term, ed hardy mens jackets
saying the Afghan Army should assume full control of the country’s security within five years. Ed Hardy chothing
“We will decrease the role of international forces,” ed hardy womens jackets
Mr. Karzai said at a ed hardy bedding
midday ceremony ed hardy hat
held at ed hardy mens jeans
the presidential ed hardy cap
palace in Kabul. ed hardy mens outerwear
“We want our ed hardy belt
security within five ed hardy womens hoodies
years to be entirely ed hardy mens hoodies
within the hands of the Afghan government ed hardy womens outerwear
and led by Afghans.”
The ceremony ed hardy jackets
was the culmination of a fraught ed hardy longsleeve
and chaotic electoral process that began on Aug. ed hardy cap
20 when Afghans went to the polls. ed hardy hat
Mr. Karzai was proclaimed ed hardy bedding
the winner earlier ed hardy belt
this month when his main challenger, christian audigier womens hoodies
Abdullah Abdullah, a former foreign minister, christian audigier hoodieswithdrew from a run-off christian audigier hoodies
after a United Nations-sponsored inquiry christian audigier womens hoodies
found evidence of widespread electoral fraud. ugg boots,
cheap ugg boots,
discount ugg boots,
australia ugg boots,
wholesale ugg boots,
sheepskin ugg boots,
ugg boots 5815,
Louie Lighting
Outdoor lighting
Chandeliers
Pendant lights
異業種交流パーティー
サイト売却
エステ 池袋
大規模修繕
ゴルフ 合コン
インターネットFAX
結婚相談所 東京
データ 通信 カード
家電 買取
英語 メール
ビーズ 作り方
アトピー 入浴
霊園 東京
葬儀 埼玉
阿比留 章雄
整体 独立
整体 スクール 東京
葬儀 横浜市
霊園 松戸
アセチルグルコサミン
目の疲れ 解消
飲む プラセンタ
口臭対策
世田谷区 矯正歯科
便秘 解消
血糖値を下げる
embassy ces
お好み焼き 渋谷
卵管閉塞 妊娠
等々力 不動産
女性 マンション
子宮腺筋症 不妊
中目黒 賃貸
都筑区 アパート
武蔵小山
事業再生
婚活
ウェディングドレス
募金
インプラント
カラコン
ボイストレーニング
債務整理
民事再生
自己破産
過払い金
任意整理
リサイクルショップ
オフィス家具
店舗デザイン
債務整理
過払い
新宿 マッサージ
タイ古式マッサージ
広島 不動産
婚活
お見合いパーティー
マカ
格安航空券 国内
バイク便
税理士 東京
青梅市 不動産
PC 移転
渋谷 賃貸
ショルダーバッグ メンズ
賃貸 恵比寿
リコメンド
看護師 求人
保育士資格
賃貸伊丹市
新聞折込
sohbet odaları
sohbet
yonja
chat siteleri
forum siteleri
toplist ekle
sohbet
yonja
netlog
sohbet
kizlarla sohbet
sohbet
sohbet
dini sohbet
islami sohbet
chat
sohbet chat
mirc indir
cinsel sohbet
porno izle
camfrog indir
lida
chat
The world's largest shoe industry portals, shoes, nets to provide a flood of Air Jordan Shoes of information, NFL Jerseys and puma chaussures hommes line trading and other information.
We brought together the world's top fashion trends of designer jeans to display their charming fashion
imagination, creative unique design, to explore the city a unique atmosphere of casual upstart. With its
fashion sense, he was ahead of the sensitive integration of the brand concept, the shackles of the
traditional dress once released, infiltrated fashion, leisure elements, breaking the traditional
framework, and vividly show the self-style, to bring traditional costumes outside the the trend of
choice. We have diesel jeans, Ed-hardy jeans, D & G jeans, Levi's jeans and other brands of jeans, first
of all to ensure adequate quality and comfort and make the product style and the beautiful, comfortable,
classic European and American avant-garde style of the performance of most vividly Cheap jeans allow you to reduce overhead at the same time leading
the fashion in our series of jeans, you can create their own unique personality delightful charm.Our
center also has NFL Jerseys, as well as a variety of clothing
cheap polo shirts
Air jordans shoes itself work very well,Wear after will feel
elasticity, very comfortable,"The function is walking shoes, so when the design will comfort in the
first,And when the sports shoes is lightsome feeling more comfortable, durability and wearing
comfortable, walk the portability of new products, etc.To what extent can actually cheaper
,Our sports Chaussures at the Sports Chaussures and Cheap Nike Shoes etc all kinds ofcheap
shoes our 2010 what you love design,our Sports Chaussures look
at the welcome your presence
Anothre set of ralph lauren polo in three colors. Ralph Lauren
Polo Shirts come with a graduation of colors on the chest. The new shiki Cheap polo shirts logo is on
the chest. Comes in all layres and can be worned tucked in or out Wholesale Polo Shirts -50% OFF.
Also comes with sculpted collar and arm cuffs.Flat knit collar, Contrast placket and half moon,
Mothre-of-pearl buttons and tennis tail, Soft double knit piqué,100% cotton ralph lauren polo shirts. If
you lend your embroidrere a sample cheapralph lauren polos that will help you get the closest match to
the original embroidreed design. The artwork you provide will sreve as a template for your embroidrey
set-up but discount ralph lauren polos can't be used to identify some stitch types, cheap polo shirts Cheap Polo Shirtsgive precise sizing of
embroidreed elements and show thread polo clothing colors.
cheap ghd
mbt
mbt sandals
mbt sale
mbt
mbt sandals
mbt sale
mbt
mbt sandals
mbt sale
mbt
mbt sandals
mbt sale
mbt
mbt sandals
mbt sale
mbt
mbt sandals
mbt sale
mbt
mbt sandals
mbt sale
mbt
mbt sandals
mbt sale
Your article is very awesome. Do you know something new about NFL Jerseys? Our Chaussure de Sport is a good store provide you high quality Nike shoes. Air Max Shoes are the best selling shoes. clothing jeans are very popular among fashion girls and boys. athletic shoes can perfectly match with it. ed hardy clothing is also very popular in recent years, Chaussures Sports will lead you a fashion life. Nike shoes like nike tn,tn requin,Tn Requin and Air Shoes are very popular among young fashion girls and boys. China Wholesale are also very cheap but functional. We also provide you top quality Tennis Racquet Shop,Cheap Nike Shoes, cheap nike shox,ed hardy, Nike Chaussures ,cheap polo shirts,Polo Shirts at a reasonable prices and awesome services.
Thanks for your great post . it help me a lot ! but I have something to share with you too ! you can login our website to look around , all our products are top quality and low prices . would you like to buy something ? ok follow me ! better choice better life ! do not leave something to regret in yourfuture!
Cheap Polo
Air Max
Nike Chaussures
Chaussures Sport
NFL Jerseys
China Wholesale
Thanks, it is very nice and helpful .i agree with you very much ! but I have something to share with you too ! you can login our website to look around , all our products are top quality and low prices . would you like to buy something ? ok follow me ! better choice better life ! do not leave something to regret in yourfuture!
NFL Jerseys
tn chaussures
chaussures puma
Tn Requin
Cheap Polo Shirts
chaussures pas cher
better choice better life !Thanks for your post , it is very nice and helpful .see you !
cheap nike shox
cheap sport shoes
nike tn dollar
ed hardy ugg boots
ed hardy love kills slowly
ed hardy clothing us
ed hardy clothing
cheap ed hardy
cheap ed hardy clothing
ed hardy clothes
ed hardy wholesale
ed hardy clothing
ed hardy t shirts
ed hardy shirts
ed hardy uk
ed hardy t shirts
ed hardy shirts
ed hardy hoodies
Cheap JORDAN SHOES,,
cheap nike max ,。
puma future cat
ed hardy ugg boots.
ed hardy love kills slowly boots.
ed hardy love kills slowly.
ed hardy polo shirts.
cheap ed hardy clothing,.
ed hardy shirts .
ed hardy t shirts.,.
artık en moda film izleme siteleri ve aşırı düşüş yaşayan sohbet sitelerine yazık oluyor biber hapı ile zayıflamak artık çok kolay biber hapı zamanı ile sizlerleyiz
kelebek mirc kelebek chat
kelebek sohbet sohbet
lannnnnnnnnn sikiş nennnnnn sikiş de porno izle izleyin quduklarım sizi :p film izle
seo mirc indir sohbet film izle haber sohbet islami sohbet
hd film izle birdeeeeee erotik film izle
Nice article and insight. Good to have bumped onto your blog and met you.
Photoshop dersleri Kitap özetleri css tutorial program indir photoshop ders ajax tutorial ilaç rehberi
film izle
Those are best online website , best service , best quality.
ed hardy clothing
Chaussures Sport
Tennis Racquet Shop
Cheap Polo Shirts
The North Face Jackets
cheap ed hardy
Chaussures Sport
Tennis Racquet
nike shox r4
ed hardy
cheap ed hardy
polo shirts
cheap polo
Remise Chaussures Sport
nike tn requin
ed hardy clothes
nike femmes chaussures
Ed hardy streak of clothing is expanded into its wholesale ED Hardy chain so that a large number of fans and users can enjoy the cheap ED Hardy Clothes range easily with the help of numerous secured websites, actually, our discount ED Hardy Outlet. As we all know, in fact Wholesale Ed Hardy,is based on the creations of the world renowned tattoo artist Don Ed Hardy. Why Ed hardy wholesale? Well, this question is bound to strike the minds of all individuals. Many people may say cheap Prada shoes is a joke, but we can give you discount Prada Sunglasses , because we have authentic Pradas bags Outlet. Almost everyone will agree that newest Pradas Purses are some of the most beautiful designer handbags ( Pradas handbags 2010) marketed today. Now we have one new product: Prada totes. The reason is simple: fashion prohibited by ugg boots, in other words, we can say it as Cheap ugg boots or Discount ugg boots. We have two kinds of fashionable boots: classic ugg boots and ugg classic tall boots. Ankh Royalty--the Cultural Revolution. Straightens out the collar, the epaulette epaulet, the Ankh Royalty Clothing two-row buckle. Would you like to wear Ankh Royalty Clothes?Now welcome to our AnkhRoyalty Outlet. And these are different products that bear the most famous names in the world of fashion, like Ankh Royalty T-Shirt, by the way-Prada, Spyder, Moncler(Moncler jackets,or you can say Moncler coats, Moncler T-shirt, Moncler vest,and you can buy them from our discount Moncler outlet), GHD, ED Hardy, designer Sunglasses, Ankh Royalty, Twisted Heart.
We provideflare True Religion Jeans of very low Price. And also Wholesale Evisu JeansYou can choose thecheap True Religion Jeansfrom our online shop. We have Clothing Jeans, mens clothing jeans, armani jeans, designer jeans,True Religion Jeans, womens jeans,skinny jeans,denim jeans in our shop.
ugg boots
polo boots
polo shoes
herve leger
herve leger handage dress
chanel outlet
chanel handbags
chanel bags
chanel iman
ralph Lauren polo
ralph lauren outlet
lacoste polo
polo raplh lauren
air jordan 2010
cheap jordan shoes
jordan ajf shoes
discount jordan shoes
jumpman23
moncler
moncler jackets
moncler coats
moncler vest
moncler outlet
moncler Polo t shirt
cheap five finger shoes
kiss ghd
We share the opinion on glasses and I really enjoy reading your article.
Good job for writing this brilliant article of eyeglasses online.
What an inspiring article you wrote! I totally like the useful eyewear info shared in the article.
This cheap eyeglasses article is definitely eye-opening and inspiring.
I greatly benefit from your articles every time I read one. Thanks for the prescription glasses info, it helps a lot.
I really like this cheap kids glasses article, and hope there can be more great resources like this.
I like your ideas about child glasses and I hope in the future there can be more bright articles like this from you.
This is the best kids sunglasses article I have ever found on the Internet.
Great article, it's helpful to me, and I also like the useful info about unisex glasses.
Excellent point here. I wish there are more and more unisex eyeglasses articles like that.
Thank you so much for sharing some great ideas of metal eyeglasses with us, they are helpful.
Bright idea, hope there can be more useful articles about plastic glasses.
I totally agree with you on the point of plastic eyewear. This is a nice article for sure.
Your do have some unique ideas here and I expect more titanium eyeglasses articles from you.
I appreciate your bright ideas in this aviator glasses article. Great work!
I love this aviator eyewear article since it is one of those which truly convey useful ideas.
I am glad to read some fantastic wood-like eyeglasses article like this.
You have given us some interesting points on rimless glasses. This is a wonderful article and surely worth reading.
It has been long before I can find some useful articles about rimless eyewear. Your views truly open my mind.
Great resources of wood-like eyewear! Thank you for sharing this with us.
Bright idea, hope there can be more useful articles about nds card.
We share the opinion on nintendo ds and I really enjoy reading your article.
I like your ideas about ez flash vi and I hope in the future there can be more bright articles like this from you.
This is the best dstti card article I have ever found on the Internet.
I love this m3 dsi article since it is one of those which truly convey useful ideas.
Thank you so much for sharing some great ideas of r4 with us, they are helpful.
I appreciate your bright ideas in this r4 dsi article. Great work!
Great article, it's helpful to me, and I also like the useful info about r4 sdhc.
I greatly benefit from your articles every time I read one. Thanks for the ak2i cards info, it helps a lot.
Your do have some unique ideas here and I expect more acekard 2i articles from you.
Excellent point here. I wish there are more and more tti upgrade articles like that.
It has been long before I can find some useful articles about nintendo ds card. Your views truly open my mind.
I really like this dstti adv price article, and hope there can be more great resources like this.
What an inspiring article you wrote! I totally like the useful DSTT cards for dsi info shared in the article.
Good job for writing this brilliant article of M3 Adapter.
This r4i gold article is definitely eye-opening and inspiring.
Great resources of R4 Revolution for DS! Thank you for sharing this with us.
I am glad to read some fantastic Acekard AK2i article like this.
You have given us some interesting points on ak2i ds lite. This is a wonderful article and surely worth reading.
I totally agree with you on the point of Acekard 2i dsi 1.4. This is a nice article for sure.
You have given us some interesting points on glasses. This is a wonderful article and surely worth reading.
I love this eyewear article since it is one of those which truly convey useful ideas.
We share the opinion on eyeglasses online and I really enjoy reading your article.
It has been long before I can find some useful articles about cheap eyeglasses. Your views truly open my mind.
Your do have some unique ideas here and I expect more prescription glasses articles from you.
Great article, it's helpful to me, and I also like the useful info about cheap kids glasses.
Thank you so much for sharing some great ideas of child eyeglasses with us, they are helpful.
Excellent point here. I wish there are more and more children eyeglasses articles like that.
Good job for writing this brilliant article of kids sunglasses .
This is the best unisex glasses article I have ever found on the Internet.
I really like this metal eyeglasses article, and hope there can be more great resources like this.
I greatly benefit from your articles every time I read one. Thanks for the plastic eyewear info, it helps a lot.
Bright idea, hope there can be more useful articles about titanium eyewear.
Great resources of aviator eyeglasses! Thank you for sharing this with us.
I am glad to read some fantastic rimless glasses article like this.
We share the opinion on nds card and I really enjoy reading your article.
It has been long before I can find some useful articles about nintendo ds. Your views truly open my mind.
Thank you so much for sharing some great ideas of ez flash vi with us, they are helpful.
This dstti card article is definitely eye-opening and inspiring.
I love this dstt card article since it is one of those which truly convey useful ideas.
I totally agree with you on the point of m3 dsi. This is a nice article for sure.
I like your ideas about r4 and I hope in the future there can be more bright articles like this from you.
I am glad to read some fantastic r4 dsi article like this.
I really like this r4 sdhc article, and hope there can be more great resources like this.
Good job for writing this brilliant article of ak2i cards.
Great article, it's helpful to me, and I also like the useful info about acekard 2i.
I greatly benefit from your articles every time I read one. Thanks for the nintendo ds card info, it helps a lot.
What an inspiring article you wrote! I totally like the useful M3 Adapter info shared in the article.
Hd Film, Video izle
Car Videos
Turkey Hotels
Wordpress Tema
Şarkı çeviri, Müzik
Film izle, Sadece Film izle
Oyun, Oyun Haberleri
Wordpress, Webmaster
Oyun indir, Oyun download
vibram kso
vibram running shoes
MBT boots
MBT shoes in fashion
cheap mbt shoes sale
discount mbt outlet 2010
MBT Walking Shoes
spyder
spyder jackets
spyder ski wear
coach outlet
coach handbag
coach handbags
coach bag
coach bags
spyder
spyder jackets
spyder ski wear
spyder
spyder jackets
spyder ski wear
ugg boots
ugg boots cheap
discount ugg boots
classic ugg boots
ugg classic tall boots
GHD Mini Styler
cheap ugg boots
discount ugg boots
ugg boots
classic ugg boots
ugg classic tall boots
Post a Comment